Introduction
Exosomes are a class of extracellular vesicles (EVs), sized around 100 nm (30–120 nm) in diameter, that are released from many different cell types. Exosomes differ from another class of EVs, microvesicles (MVs, also referred to as 'ectosomes') that are of a different size (up to 2 µm) and are produced by a different mechanism. Exosomes were first discovered over 30 years ago and were considered to be little more than a means of cellular garbage disposal. However, more recent studies have provided scientific evidence that exosomes play roles in intercellular communication. Advances in the study of exosomes have identified the mechanisms regulating their production and have shown that exosomes contain cell-type specific collections of proteins, lipids and genetic material that can be transferred to recipient cells where they regulate function. Recently, scientific attention has focused on the various functions of exosomes in the pathophysiology of diseases including cancer, metabolic diseases, cardiovascular diseases, immune diseases, and neurodegenerative diseases. In particular, it has been shown that in type 2 diabetes mellitus (T2DM), exosomes secreted from skeletal muscle, visceral adipose tissue (VAT) and hepatocytes can transfer both functional proteins and RNA species that regulate the metabolic function of both remote tissues and of adjacent cells1,2,3).
The range of biomolecules in exosomes is diverse, including soluble and membrane-bound proteins, lipids, mRNA, microRNAs (miRNAs) and noncoding RNAs (ncRNAs). Therefore, exosomes are potentially a good source of disease biomarkers for early diagnosis and/or prediction of disease progression. However, there are several unresolved problems with the isolation of exosomes from biological fluids. In this review, we describe the potential roles of exosomes from diverse cell types in the regulation of cell metabolism, and discuss the possibility of the clinical use of blood-based exosomal biomarkers and the challenges for the use of exosomes as a source of disease biomarkers.
Production of exosomes and exosome cargos that regulate metabolic function
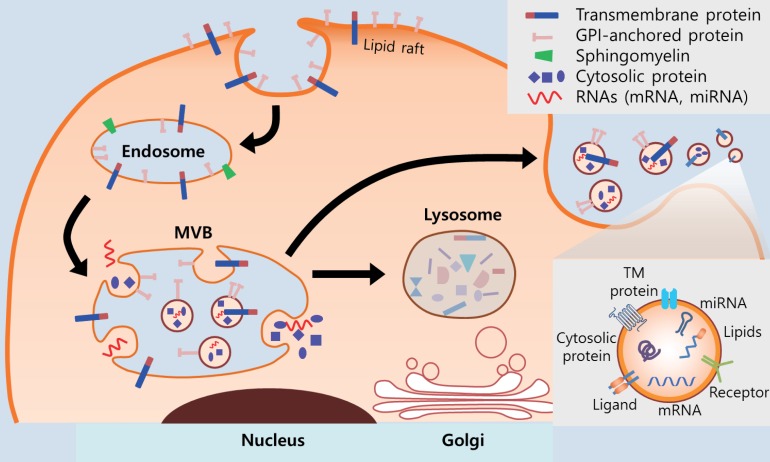
Exosomes originate from sections of the plasma membrane that are heavily enriched in phosphatidylserine. As illustrated in Fig. 1, the first step of exosome formation is inward budding of the plasma membrane to form an early endosome. During maturation of the endosome, the inward budding of limited areas of the endosomal membrane to form nano-sized vesicles produces multivesicular bodies (MVBs) that contain intraluminal vesicles (ILVs), in which cytoplasmic components including soluble proteins and nucleic acids are present. Although the mechanisms by which proteins are loaded into exosomes are not fully understood, the endosomal sorting complexes required for transport machinery is essential for the formation of ILVs in an MVB4,5). The inward budding of ILVs and the sorting of cytosolic proteins or nucleic acids also involve tetraspanin- and ceramide-dependent interactions6). Once MVBs are formed, the ILVs (exosomes) are released to the extracellular environment by fusion of the MVBs with the plasma membrane, a process that is regulated by a small GTPase-dependent mechanism. Alternatively, MVBs can fuse with a lysosome or autophagosome, where the contents of the MVBs are degraded (Fig. 1). The secreted exosomes function in fundamental pleiotropic biologic processes via direct interaction of their transmembrane proteins or lipid ligands with cell surface receptors, or by delivering their contents (e.g., transcription factors, cytosolic proteins, mRNA, miRNA and ncRNA) into recipient cells via membrane fusion7). Their biologic effects can be local (such as regulation of synaptic plasticity) or effected by delivery of secreted exosomes to remote target cells. These physiologic effects include, but are not limited to, the regulation of immune and neuronal function8,9), cell phenotype modulation10,11) and tissue regeneration12,13). It is not surprising that exosomes also have significant pathological roles in a variety of diseases including tumor progression14,15), promoting immune escape16), transmission of neuropathological proteins17,18), and metabolic dysfunction19).
Although studies that clearly demonstrate the nature of EVs are limited, the number of EVs is increased in the plasma of patients with insulin resistance and T2DM20,21). In particular, lipids or surface molecules of EVs released from skeletal muscle, platelets, or T cells increase the risk of metabolic dysfunction or atherogenic tendency19,20,21). In addition, an increased number of EVs has also been linked with obesity, micro/macrovascular complications, and inflammatory conditions22,23,24,25), indicating that the quantity of exosomes can be an indicator of these diseases. Recent studies demonstrated that exosomes can orchestrate insulin signaling in peripheral tissues, which suggested the emerging concept of exosome-mediated regulation of metabolism in T2DM19,26,27,28,29). For example, exosomes released from VAT in diabetic mice can fuse with blood monocytes and stimulate their differentiation into activated macrophages30). Exosomes released from VAT in mice with T2DM also induced the development of insulin resistance when they were administered to normal mice30). Adipocyte-derived exosomes contain hundreds of proteins including adipokines28), which may indicate a role for exosomes in the intercellular transmission of signals. Exosomes released from skeletal muscle, another critical tissue regulating whole-body metabolism, can be incorporated into various tissues including skeletal muscle, pancreas and liver, and can modify muscle homeostasis19).
Based on these findings and advances in other diseases, attempts are being made to develop novel diagnostic or prognostic biomarkers using exosomal molecules isolated from biological fluids. For example, the presence of diseaseor stage-specific miRNAs in blood exosomes may be a reliable and clinically applicable candidate biomarker for diagnosis or prediction of various diabetic complications31,32,33).
Exosomes in biological fluids
A biomarker is defined as 'a characteristic that is objectively measured and evaluated as an indicator of a normal biologic process, pathogenic process, or pharmacologic response to a therapeutic intervention'34). Exosomes can be detected in a variety of biological fluids including serum, plasma, urine, saliva, cerebrospinal fluid (CSF) and milk. In particular, serumor plasma-derived exosomes are a promising avenue to be explored for biomarker development. T2DM is the principal component of metabolic syndrome (MetS); therefore, based on the known pathophysiologic processes of MetS, blood-based biomarkers of T2DM could offer a minimally invasive method to predict patients' outcomes, a modality for predicting the response to therapeutic intervention, and an opportunity to provide earlier care to avoid the development of more severe complications. Patients with different types of diabetes and different complications of diabetes showed diverse patterns of exosomal biomolecules, suggesting that exosomes may contribute to tissue- or stage-specific pathogenic mechanisms of diabetes and its complications35). Exosomes from specific tissues including liver, muscle, endothelium, blood cells and VAT can travel in the bloodstream and be distributed to remote tissues. In the target tissues, exosomes that are derived from remote or adjacent tissues integrate with or interact with recipient cells, and act to regulate gene transcription, posttranslational modification, signal transduction, and metabolism. In fact, white VAT, an important organ that regulates whole-body metabolism and metabolic signaling and modulates the function of remote tissues including liver36) and macrophages30), is known to release exosomes. Notably, VAT dysfunction is an independent risk factor for cardiovascular complications of T2DM37,38). The pathophysiological mechanisms that link cardiovascular complications and VAT dysfunction are still poorly understood, but blood-based biomarkers that indicated the risk of diabetes complications would be extremely useful. Because diverse cellular cargoes are delivered by exosomes, these could be a source of blood-based biomarkers. Therefore, exosomes isolated from plasma or serum might present a molecular signature of a combination of tissue types, and could represent both common and specific markers of diverse cells and tissues or disease status. In contrast, exosomes from other types of biological fluids such as CSF, milk, or saliva may be tissue specific. The matrix of the biological fluid may also influence the exosome enrichment procedure; for example, albumin in plasma can be a major protein contaminant of isolated exosomes. Therefore, the procedure by which exosomes are isolated from biological fluids is critical for the development of exosomal biomarkers using various types of biological fluid.
Issues with exosome purification for biomarker development
The characterization of samples for exosome enrichment is the initial and essential step for exosomal biomarker development, because the extracellular milieu is complex, and plasma and serum, in particular, are biological fluids that reflect the extracellular environment of entire organs and of endothelium and blood cells. Therefore, the efficiency and recovery of exosomes isolated from biological fluids and the contaminants in the isolated exosomes are dependent on the matrix of the sample. There are several proposed methods to enrich exosomes from biological fluids and in vitro cell culture medium, but the optimal method of exosome isolation has not been established. The most widely used method for exosome enrichment is differential centrifugation, in which sequential rounds of centrifugation are intended to pellet cellular debris (10 minutes at 300 g), large vesicles (10 minutes at 2,000 g), MVs larger than 150 nm (30 minutes at 10,000 g) and exosomes (ultracentrifugation for 70 minutes at 100,000 g). Washing the final pellet that is enriched with exosomes can reduce its contamination by soluble proteins.
However, there are several issues with standardized exosome isolation using the differential centrifugation method. First, the duration of each centrifugation must be optimized according to the rotor type used. Two types of rotors are commonly used: fixed-angle or swing-bucket rotors. Because the sedimentation efficiency of EVs of diverse size and density is dependent on the sedimentation path length, the duration of centrifugation must be calculated for each rotor type39). Second, contamination of exosomes prepared by ultracentrifugation with small nonexosomal vesicles must be considered. Indeed, MVs that are generated by blebbing from plasma membranes have a broad size range (50–2,000 nm) that overlaps with the size of exosomes40,41). Therefore, we cannot exclude the possibility that a disease-specific exosomal biomarker is actually a combination of biomolecules from exosomes and small MVs. To exclude this possibility, in addition to size-based characterization the isolated exosomes should be characterized using at least three exosomal markers (e.g., tetraspanins, TSG101, and Alix) and the absence of or low levels of contamination with nonexosomal markers (e.g., Grp94, GM130, and cytochrome C) should be confirmed42). Exosomes can be further characterized using a combination of immunodetection of exosomal markers with electron microscopy (EM). EM is valuable for assessments of the morphology and size of vesicles in the samples (Fig. 2). Third, ultracentrifugation can induce the collapse and damage of exosomal membranes or aggregation of exosomes. Damage to the exosomal membrane can influence the quantification of biomarker levels. The formation of aggregates composed of a mixture of EVs of various phenotypes and morphologies may lead to misinterpretation of the level of exosomal biomarkers43). Finally, the conditions for differential centrifugation should be optimized according to the specific characteristics of each biological fluid (e.g., the difference in viscosity between plasma and urine).
Exosomes can also be purified by density gradient (DG) isolation with iodixanol (OptiPrep, Sigam-Aldrich, St. Louis, MO, USA) using a sucrose gradient followed by ultracentrifugation. Ultracentrifugation of samples overlaid on an iodixanol gradient for 16 hours at 100,000 g produces a layer that contains exosomes, which corresponds to a buoyant density of 1.13–1.19 g/mL. This method can prepare a relatively homogeneous exosome fraction with low levels of contamination44,45), and may be the best method for preparing highly purified exosomes for in vitro experiments. However, the preparation of exosomes by DG is relatively labor intensive, poorly reproducible and hard to standardize for routine clinical application.
Another method to prepare exosomes is size-exclusion chromatography (SEC). One or several fractions of a sample are loaded onto a packed SEC column that allows passage of nondamaged vesicles of regular shape and homogeneous size46). Larger vesicles can enter fewer pores in the column than can the smaller vesicles. The main advantages of this procedure for clinical applicability are its simplicity, good reproducibility and minimal damage to vesicles. However, this method cannot exclude the possibility of protein or RNA contamination, and because the sample volume should not exceed 10% of the resin volume the SEC method is less useful for larger-volume samples (e.g., urine or cell culture media) because of the cost. This limitation can be partially resolved by combining SEC with the use of commercially available ultrafiltration units (e.g., Vivaspin [Sartorius, Bohemia, NY, USA] or Amicon [Millipore, Darmstadt, Germany]) that keep exosomes on the filter surface and allow the removal of the majority of the sample fluid.
Finally, commercially available kits for polymer-based precipitation or immune bead-based capture methods (e.g., ExoQuick [System Biosciences, Palo Alto, CA, USA], Exo-spin [Cell Guidance Systems, St. Louis, MO, USA]) have been used in several biomarker studies to isolate exosomes from plasma47,48). The polymer-based precipitation kits enable high-throughput, fast and simple isolation of exosomes from low-volume samples. However, it should be noted that the purity of the exosomes was inferior to that obtained by other methods49) and that contamination by extracellular proteins was a major concern. The immune bead-based capture method produces a highly selected subpopulation of vesicles that is specific to a certain population or disease status and may not be generalizable. In addition, the availability of suitable antibodies for exosome isolation may be a limiting factor.
Future perspectives for exosomal biomarkers
Compared with healthy controls, there were significantly more plasma EVs in patients with MetS50,51), supporting the hypothesis that quantification of EVs in blood may have potential as a screening biomarker for MetS. There are limited data on the clinical applicability of exosomal biomarkers for prediction of T2DM progression. However, considering the biologic roles of the molecular contents of exosomes in regulation of tissue-specific and/or whole-body metabolism, several studies have supported the hypothesis that proteins or miRNA in circulating or urinary exosomes (or microparticles) may be potential biomarkers to predict disease progression. For example, certain proteins and miRNAs in circulating or urinary EVs are associated with cardiometabolic risk factors21,23,29,52,53,54) and renal complications of diabetes55,56,57), respectively. In addition, these exosomal biomarkers may be useful for monitoring the pharmacological or therapeutic responses of patients with MetS and T2DM. For example, the levels of miRNAs let-7a and let-7f in circulating EVs were normalized by oral antidiabetes medication29), although it is unclear whether these changes were the result of pharmacological regulation of the underlying pathophysiology or were merely related to an improvement of glucose control.
These clinical studies provided molecular evidence that exosomes in biological fluids are good sources of potential biomarkers for metabolic diseases. There are currently no biomarkers of circulating or urinary exosomes that are validated for use in routine clinical practice. Nevertheless, although the analysis of circulating or urinary exosomes in metabolic disease is currently limited, exosomes may be a good source of diagnostic and prognostic biomarkers. To this end, longitudinal clinical studies with large sample size and with standardized system for the analysis of exosomal cargos should be conducted to evaluate clinical values of exosomal biomarkers for prediction of the progression of metabolic diseases.
In conclusion, there is increasing evidence that exosomes play roles in the regulation of tissue-specific and/or whole-body metabolism via the delivery of diverse exosomal cargo molecules to adjacent and remote tissues. Recent advances in the understanding of the biologic functions of exosomes facilitate clinical research towards the identification of biomarkers by using several methods to isolate exosomes from biological fluids. However, global efforts towards the standardization of preanalytical factors and procedures for exosome isolation (e.g., by the International Society for Extracellular Vesicles) are critical to facilitate the identification of clinically applicable exosomal biomarkers and overcome current challenges58). In addition, the characteristics of specific biological fluids used for exosome isolation must be considered (e.g., the effects of the different viscosities and volumes of plasma and urine samples). The development of a less labor intensive, cheap, high-throughput and valid method for purifying exosomes from biological fluids is a prerequisite for the identification of clinically applicable exosomal biomarkers.